Строение клеточной стенки грибов
Клетка гриба состоит из клеточной оболочки (снаружи она часто бывает покрыта слизистым слоем — капсулой), ломасом, цитоплазмыс цитоплазматической мембраной, эндоплазматнческой сетью, митохондриями, рибосомами, аппаратом Гольджи (диктиосомами) и ядрами (рис. 2). Иногда в клетке грибов есть вакуоли и различные включения.

Рисунок 2 – Строение клетки гриба
Клеточная оболочка, осуществляющая у грибов многочисленные функции, в том числе активного всасывания питательных веществ из субстрата, в качестве основных компонентов содержит хитин, полисахариды, в том числе глюканы, белки и жиры. Обычно слой молекул глюканов покрывает сверху молекулы хитина. Хитин — вещество, содержащее азот,— в оболочках многих видов грибов составляет до 60% сухой массы. Химическая природа хитина — линейный полимер, в молекулу которого входят ß-1,4-связанные единицы N-ацетилглюкозамина.
В оболочках грибных клеток они собраны в кристаллиты.
лисахариды (d-глюкоза, N-ацетилглюкозамин, d-манноза) составляют до 80—90% содержимого оболочки клетки. В клеточной оболочке грибов имеются также пигменты (меланины, хиионы), сюда же входят различные ионы и соли. Электронно-микроскопическое изучение оболочек клеток грибов показывает, что они состоят из нескольких слоев фибриллярного строения. Эти фибриллы, представляющие собой белковые микротрубочки, образуют скелет, который служит основой для остальных компонентов оболочки. Клеточная оболочка придает форму клеткам гиф и органам размножения. Ее поверхность является местом локализации некоторых ферментов. Проницаемость клетки и ее способность связывать определенные вещества играют роль в питании грибов, их отношении к фунгицидам, антибиотикам.
В результате лизиса клеточная оболочка грибов может разрушаться двумя путями: воздействием ферментов, выделяемых другими клетками или организмами, и воздействием ферментов, образуемых в клетке самого гриба. Наиболее важными ферментами, катализирующими разложение отдельных компонентов клеточной оболочки грибов, являются α- и ß-глюканазы, протеазы и пептидазы, целлюлазы, липазы, хитиназы, гексозаминидазы, глюкуронидазы, глюкозамннидазы, целлобиазы. Комплекс ферментов зависит от особенностей компонентов клеточной оболочки разных видов грибов. На поверхности клеточной оболочки и цитоплазматической мембраны локализованы ферменты, превращающие не усваиваемые клеткой полимеры в усваиваемые мономеры. Отличительным признаком клеточной оболочки некоторых представителей низших грибов является отсутствие в ней хитина и наличие только целлюлозы.
Источник: studopedia.su
Общее строение грибов
По своей биологической сути грибы – многоклеточные организмы. Каждая клетка гриба, как и бактериальная, животная и растительная, нуждается в энергии и в определенном объеме биологически активных веществ. По общим морфологическим признакам строение клетки гриба очень похоже на строение животной и растительной клеток, так как грибы – эукариоты. Грибная клетка имеет ядро в центре, которое выполняет стандартные для себя функции – хранение генетической информации и синтез РНК. Практически наполовину клетка гриба заполнена цитоплазмой, которая содержит в своем строении основные органеллы клетки. Найдены в клетке гриба и лизосомы, содержащие различные ферменты, благодаря которым они расщепляют органические вещества. Грибная клетка также имеет и полноценный аппарат Гольджи, как у растений и у животных. Более подробно о строении животной клетки можно почитать тут.
Особенностью в строении грибной клетки является специфическая клеточная стенка, образованная хитином. Интересно, что у каждого царства животных она образовывается разными веществами. У растений – целлюлозой, у бактерий – пектином, у грибов – хитином, а у животных она вообще отсутствует. У грибов клеточная стенка берет участие в регуляции воды и газов в клетке. Как и у всех эукариот, грибная клетка имеет в своем строении рибосомы и митохондрии. В рибосомах происходит синтез белков, а митохондрии участвуют в цикле превращения энергии.
Грибы имеют цитоскелет – это основная опорно-двигательная система клетки. Есть в наличии и хромосомы с вакуолями. Хромосомы содержат ДНК, а вакуоли способны накапливать в своем теле различные клеточные соки, чем помогают успешному протеканию всех биохимических процессов в клетке. Абсолютно во всех клетках живых организмов есть клеточная мембрана, которая выполняет ряд важных функций. И грибы не исключение. В строении клетки гриба клеточная мембрана выполняет транспортную, механическую, барьерную, рецепторную, энергетическую и другие функции.
Источник: life-students.ru
Клеточные стенки высших растений
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жесткость клетки для структурной и механической поддержки, придают форму клетке, направление ее роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургора, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают против патогенов, проникающих из окружающей среды, и запасают углеводы для растения. Растительные клеточные стенки строятся прежде всего с углеводного полимера целлюлозы.
В течение цитокинеза сначала с клеточной пластинки формируется промежуточная ламела, и первичная клеточная стенка расширяется внутри промежуточной ламели.
ктическая структура клеточной стенки не всегда четко определена, существуют несколько моделей ее структуры: модель перекрестных ковелентних связей, модель привязи, модель диффузного слоя и модель стратифицированного слоя. Однако, первичная клеточная стенка может быть определена как сеть волокон целлюлозы направленных в случайных направлениях. Волокна воздержатся вместе водородными связями, обеспечивает достаточно высокую прочность. Клетки держатся вместе и разделяют желатиновую мембрану, называет его промежуточная ламела, которая содержит пектаты (соли пектиновой кислоты) магния и кальция. Клетки взаимодействуют через плазмодесмы, то есть цитоплазмени канальцы, соединяющие цитоплазмы клеток через клеточные стенки.
В некоторых типах клеток некоторых растений, после того как достигается максимальный размер или определенная точка развития, между растительным клеткой и первичной стенкой формируется вторичная стенка. В отличие от первичной стенки, волокна выравниваются в основном в одном направлении, и с каждым дополнительным слоем их ориентация слегка меняется. Клетки с вторичными клеточными стенками очень жесткие. Межклеточное взаимодействие все еще возможна через отверстия во вторичной клеточной стенке, позволяющие плазмодкрми проникать через обе клеточные стенки.
Основные углеводы, составляющих первичную клеточную стенку, это целлюлоза, гемицеллюлоза и пектин. Целлюлозные микроволокна связываются через хемицелюлозни мостики, формируя целлюлозно-хемицелюлозну сеть, которая окружена матрицей пектина. Самый общий тип хемицелюлозы в первичной клеточной стенке — ксилоглюкан.
Растительные клеточные стенки также содержат ряд белков, распространенные включают богатые гидропролин гликопротеины (HRGP), также известные как екстенсины, арабиногалактан протеины (AGP), богатые глицин протеины (GRP) и богатые пролин протеины (PRP). За исключением богатых глицин протеинов, все эти белки гликозилированные и содержат гидроксипролин (Hyp). Каждый класс гликопротеинов определяется характерной, часто повторяющейся последовательности аминокислот. Причудливые протеины содержат два или более областей, каждая из последовательностью от другого класса гликопротеинов. Большинство белков клеточной стенки перекрестных связаны с углеводами клеточной стенки и, возможно, имеют структурные функции.
Вторичные клеточные стенки могут содержать лигнин и суберин, делая стенки твердыми. Относительный состав углеводов, дополнительных компонентов и белков зависит от вида растения, типа и возраста клеток.
Клеточные стенки водорослей
Подобно высших растений, большинство водорослей, независимо от происхождения, имеют клеточные стенки. Клеточные стенки водорослей содержат целлюлозу и многие другие гликопротеинов. Дополнительные полисахариды в клеточных стенках водорослей раньше часто использовали как один из признаков для установления их таксономии, но некоторые несвязанные группы имеют те же полисахариды.
- Манозилови микроволокна зучтричаються в клеточных стенках многих зеленых водорослей (примитивних растений), в том числе родов Codium, Dasycladus и Acetabularia, и некоторых красных водорослей например, родов Porphyra и Bangia.
- Ксилан
- Альгиновая кислота — обычный полисахарид в клеточных стенках бурых водорослей
- Сульфоновани ролисахариды встречаются клеточных стенках большинства водорослей, например, в красных водорослях находим агарозу, каррагинан, порфиран, фурселеран и фуноран.
Другие вещества, которые могут быть найдены в водорослевых клеточных стенках, включают спорополенин и кальций.
Группа диатомовых водорослей (диатомей) синтезируют клеточные стенки (также известно как фрустулы или клапаны) с Кремнев кислоты (особенно ортокремневои кислоты, H 4 SiO 4). Кислота полимеризуется внутриклеточно, затем стенка вытесняется наружу для защиты клетки. Такие стенки требуют меньше энергии на свой синтез, позволяя сохранить много энергии, что, возможно, частично отвечает за быстрый рост этих организмов.
Клеточные стенки грибов
Не все виды грибов имеют клеточные стенки, но если стенка присутствует, она состоит из глюкозамина и хитина, того же углерода, что придает прочность экзоскелета насекомых. Они служат подобной цели, что и клеточные стенки растений, предоставляя грибным клеткам твердость и поддерживать свою форму и предотвращения осмотическому лизису. Они также ограничивают вход молекул, потенциально ядовитых для гриба, например некоторым фунгициды растительного и искусственного происхождения. Состав, свойства и форма грибной клеточной стенки меняются на протяжении жизненного цикла и зависят от условий произрастания.
Другие эукариоты
Оомицеты, патогены растений похожи на грибы, также имеют целлюлозные клеточные стенки. До недавнего времени за этот признак их относили к грибам, структурные и молекулярные свидетельства заставили отнести их к гетероконтив, как и бурые водоросли.
Клеточные стенки бактерий
Как в других организмах, бактериальные клеточные стенки обеспечивают структурную целостность клеткам. У прокариот, первичная функция клеточной стенки — защитить ячейку от внутреннего тургора, вызванным высокой концентрацией белков и инших молекулы внутри клетки по сравнению с окружающей средой. Бактериальная клеточная стенка отличается от других тем, что ее основным компонентом является пептидогликан, слой которого размещается немедленно вне цитоплазматической мембраны. Пептидогликан отвечает за твердость стенки и придания формы клетке. Он относительно пористый и не мешает потока растворимых молекул сквозь него. Существует два основных типа бактериальных клеточных стенок, по этому признаку бактерии делятся на грам-отрицательные и грам-положительные.
Грам-положительные бактерии
Грам-положительные бактерии характеризуется присутствием очень толстого слоя пептидогликана, который отвечает за содержание красителя кристал-виолет течение процедуры окрашивания по Граму. Такая стенка находится исключительно в организмах, принадлежащих к типам Actinobacteria (или грамм-положительные организмы с высоким содержанием G + C) и Firmicutes (или грамм-положительные организмы с низким содержанием G + C). Бактерии в пределах типа Deinococcus-Thermus также могут положительно краситься по Граму, но содержат некоторые структуры клеточной стенки, типичные от грамотрицательных организмов. В клеточную стенку грамположительных бактерий встроенные полимерного спирты, тейхоевая кислота, некоторые из которых связываются с липидами, формируя липотейхоеву кислоту. Ци вещества отвечают за соединение пептидогликана с цитоплазматической мембраной. Тейхоевая кислота придает клетке отрицательный электрический заряд благодаря наличию фосфодиестерних связей между мономерами тейхоевая кислоты.
Грамотрицательные бактерии
В отличие от грамположительных бактерий, грамотрицательные бактерии содержат очень тонкий слой пептидогликана, отвечающий за неспособность клеточных стенок содержать краситель кристал-виолет течение процедуры окрашивания по Граму. В дополнение к слою пептидогликанов, грамотрицательные бактерии имеют вторую, так называемую внешнюю мембрану, находится кнаружи от клеточной стенки и компонует фосфолипиды и липополисахариды на своей внешней стороне. Отрицательно заряженные липополисахариды также предоставляют клетке отрицательный электрический заряд. Химическая структура липополисахаридив внешней мембраны часто уникальная для отдельных штаммов бактерий и часто отвечает за реакцию антигенов с представителями этих штаммов.
Как любой двойной слой фосфолипидов, внешняя мембрана достаточно непроницаема для всех заряженных молекул. Однако, белковые каналы (погрузится) присутствуют во внешней мембране, позволяют пассивный транспорт многих ионов, сахара и аминокислот через внешнюю мембрану. Таким образом, эти молекулы присутствуют в периплазматическое, слое между внешней и цитоплазматической мембранами. Периплазматическое содержит слой пептидогликана и много белков, шо отвечают за гидролиз и прием внеклеточных сигналов. Читается, что перивлазма гелеобразная, а не жидкая, из-за высокого содержания белка и пептидогликана. Сигналы и живильни вещества с периплазматическое попадают в цитоплазму клетки используя транспортные белки в цитоплизматичний мембране.
Клеточные стенки архей
Хотя они и не уникальны, клеточные стенки архей несколько отличаются от бактериальных. Например, клеточные стенки большинства архей образованные внешними слоями белков или S-слоем. S-слоя распространенные в бактериях, где они служат единственным компонентом клеточной стенки в некоторых организмах (например в Planctomyces) или внешним слоем во многих организмах с пептидогликана. За исключением одной группы метаногенов, археи не имеют пептидогликаннои стенки. Даже в данном случае, пептидогликаны очень отличается от типа, найденного в бактериях.
Источник: info-farm.ru
Полисахариды клеточных стенок грибов, в том числе дрожжей, представлены в основном гемицеллюлозами: глюканами, хитином и маннанами. Глюканы — продукты конденсации глюкозы с разным числом звеньев —принадлежат в основном к b-1,4- и b-1,6-типу. Эти гомополисахариды формируют в клеточной стенке грибов микрофибриллярный скелет, подобный тому, который создает целлюлоза в клеточных стенках растений.
Хитин представляет собой гомополисахарид, образованный цепями N-ацетилглюкозамина, который также является структурной частью муреина (рис. 5.4). Остатки N-ацетилглюкозамина в хитине связаны между собой с помощью b(1®4)-гликозидных связей и формируют слоистую структуру, подобную целлюлозе. Однако в хитине межцепочечные водородные связи оказываются более прочными, поскольку в их образовании принимают участие N-ацетильные группы. Хитин служит также основным структурным полисахаридом покровов тела насекомых.
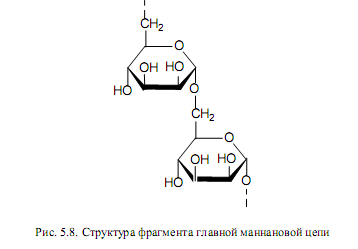
Маннаны являются полимерами маннозы (рис. 5.8). В них от главной a-1,6-цепи отходят короткие ветви (из 1—3 маннозных звена), присоединенные через a(1→2)- и a(М13)-гликозидные связи. Маннаны и глюканы — это основные полисахариды клеточных стенок дрожжей (составляют в них 60—80% сухого вещества).
Кроме гемицеллюлоз, в состав клеточных стенок грибов входят белки (6— 13%), липиды (2—9%) и неорганические полифосфаты. Белки богаты серусодержащими аминокислотами и присутствуют в виде комплексов с полисахаридами. Наиболее часто среди таких комплексов встречаются маннанопротеины. Кроме структурной функции, маннанопротеинам принадлежит рецепторная: они воспринимают сигналы из внешней среды и участвуют в осуществлении клеточных контактов. Структура маннанопротеинов определяет иммунологические свойства дрожжей.
Анализ наиболее распространенных клеточных полисахаридов, охарактеризованных в этой главе, позволяет заключить, что их структура и функции тесно связаны: полисахариды с рыхлой, разветвленной, легко доступной ферментам структурой выполняют в клетках резервную функцию. Наоборот, полисахариды, призванные служить механической опорой и защитой для клеток, имеют сложную, компактную структуру, которая упрочняется при взаимодействии полисахаридов с веществами других классов: белками, липидами, тейхоевыми кислотами и др. В этих комплексах практически всегда присутствуют межмолекулярные связи, что делает клеточные стенки жесткими, прочными, трудно доступными ферментативному расщеплению.
Источник: helpiks.org