Пассивный хроматин
Термином «хроматин» обозначают комплекс ядерной ДНК с белками (гистоны, негистоновые). Различают гетеро- и эухроматин.
Гетерохроматин. Гетерохроматин — транскрипционно неактивный, конденсированный хроматин интерфазного ядра. В СМ — базофильные глыбки. в ЭМ —скопления плотных гранул Располагается преимущественно по периферии ядра и вокруг ядрышек. Типичный пример гетерохроматина — тельце Барра.
Эухроматин. Эухроматин — транскрипционно активная и менее конденсированная часть хроматина, локализуется в более светлых участках ядра между гетерохроматином.
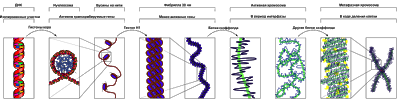
Наблюдения за структурой хроматина с помощью электронного мик-роскопа показали, что как в препаратах выделенного интерфазного хроматина или выделенных митотических хромосом, так и в составе ядра на ультратонких срезах всегда видны элементарные хромосомные фибриллы толщиной 20—25 нм.
В химическом отношении фибриллы хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов (ДНП), в состав которых входят ДНК и специальные хромосомные белки — гистоновые и негистоновые.
составе хроматина обнаруживается также РНК. Количественные отношения ДНК, белка и РНК составляют 1:1,3:0,2. Обнаружено, что длина индивидуальных линейных молекул ДНК может достигнуть сотен микрометров и даже сантиметров. Среди хромосом человека самая большая первая хромосома содержит ДНК с общей длиной до 4 см. Суммарная длина молекул ДНК во всех хромосомах одной клетки человека составляет около 170 см, что соответствует 6 • 10~12 г.
В хромосомах существует множество мест независимой репликации, т.е. удвоения ДНК — репликонов. ДНК эукариотических хромосом представляют собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. В составе генома человека должно встречаться более 50 000 репликонов, участков ДНК, которые синтезируются как независимые единицы.
Синтез ДНК как на участках отдельной хромосомы, так и среди разных хромосом идет неодновременно, асинхронно. Так, например, в некоторых хромосомах человека репликация наиболее интенсивно начинается в плечах хромосом и заканчивается (при высокой интенсивности включения метки) в центромерном районе (см. ниже). Наиболее поздно репликация заканчивается в хромосомах или в их участках, находящихся в компактном, конденсированном состоянии. Таким примером может являться поздняя репликация генетически инактивированной Х-хромосомы у женщин, формирующей в ядре компактное тельце полового хроматина.
Белки хроматина составляют 60—70 % от его сухой массы. К ним относятся так называемые гистоны и негистоновые белки. Негистоновые белки составляют 20 % от количества гистонов. Гистоны — щелочные белки, обогащенные основными аминокислотами (главным образом лизином и аргинином). Очевидна структурная роль гистонов, которые не только обеспечивают специфическую укладку хромосомной ДНК, но и имеют значение в регуляции транскрипции. Гистоны расположены по длине молекулы ДНК не равномерно, а в виде блоков. В один такой блок входят 8 молекул гистонов, образуя так называемую нуклеосому. Размер нуклеосомы около 10 нм.
При образовании нуклеосом происходит компактизация, сверхспирализация ДНК, что приводит к укорачиванию длины хромосомной фибриллы примерно в 7 раз. Сама же хромосомная фибрилла имеет вид нитки бус или четок, где каждая бусина — нуклеосома. Такие фибриллы толщиной 10 нм дополнительно продольно конденсируются и образуют основную элементарную фибриллу хроматина толщиной 25 нм.
В интерфазе фибриллы хроматина образуют петли. Эти петли собраны в розетки, где основания нескольких петель связаны друг с другом негистоновыми белками ядерного матрикса. Такие петлевые группы (петлевые домены) при падении активности хроматина могут конденсироваться, уплотняться, образуя так называемые хромомеры или хромоцентры интерфазных ядер. Хромомеры выявляются также в составе митотических хромосом.
В ядрах, кроме хроматиновых участков и матрикса, обнаруживаются
перихроматиновые фибриллы, перихроматиновые и интерхроматиновые гранулы. Они содержат РНК и встречаются практически во всех активных ядрах, представляют собой информационные РНК, связанные с белками, — рибонуклеопротеиды (информосомы). Матрицами для синтеза этих РНК являются разные гены, разбросанные по де конденсированным участкам хромосомных (хроматиновых) фибрилл.
Особый тип матричной ДНК, а именно ДНК для синтеза рибосомной РНК, собран обычно в нескольких компактных участках, входящих в состав ядрышек интерфазных ядер.
ХРОМОСОМЫ.
Хромосомы видны полностью только в митозе. Наиболее удобно изучать их и метафазе (метафазные хромосомные пластинки). Основными химическими элементами хромосомы являются ДНК и белки. Комплекс ДНК с белками (в основном с гистонами) формирует фибриллярную структуру — элементарную хромосомную фибриллу, имеющую нуклеосомную организацию. Каждая нуклеосома представляет собой комплекс из 8 молекул гистонов (гистоновый октамер). Вокруг него молекула ДНК образует около 2 оборотов. Участки ДНК, связывающие соседние нуклеосомы, называются линкерной ДНК. Следующим уровнем организации хромосомы является нуклеомерная организация, или уровень хроматиновой фибриллы. В ней нуклеосомы объединяются в нуклеомеры, причем каждый нуклеомер состоит из 8—10 нуклеосом и имеет диаметр около 30 им. Хромосомы образованы хроматиноными фибриллами (хроматидами) в интерфазе. В ходе последующей упаковки нуклеомер подвергается суперспирализации и превращается в хрономер, содержащий петельные домены. Петельный домен имеет диаметр до 300 нм и соответствует одному или нескольким генам. Хромомер далее за счет суперспирализации
укорачивается, образуются конденсированные хромосомы, видимые только в митозе клетки.
МОРФОЛОГИЯ И КЛАССИФИКАЦИЯ ХРОМОСОМ
Каждая хромосома в световой микроскоп выглядит в виде палочки. У большинства хромосом можно увидеть первичную перетяжка — центромер, или кинетохор. Он делит хромосомы па два плеча. Если длина плеч одинакова, такие хромосомы называются метацентрическими. Если одно плечо больше, то хромосомы являются субметацентрическими. Хромосомы с очень коротким одним плечом называются акроцентрическими. Иногда на хромосомах есть вторичные перетяжки, отделяющие от хромосомы маленький участок — сателлит. В области вторичных перетяжек находятся ядрышковые организаторы.
Согласно Денверской классификации хромосом (Денвер, США. 1960), учитывающей размеры хромосом, расположение первичных, вторичных перетяжек и наличие спутника, все хромосомы делятся на 7 групп (А, В, С, D, E, F, G). Существует также Парижская (1971) классификация хромосом. В ее основу положена дифференциальная окраска хромосом некоторыми красителями. Эта окраска выявляет в хромосомах чередующиеся светлые и темные полосы (гетеро- и эухроматиновые районы), уникальной для каждой пары хромосом. Дифференциальная окраска позволяет достоверно отличить одну пару хромосом от другой.
12. Основные положения клеточной теории. Определение клетки. Понятие о жизненном цикле клеток: его этапы и их морфофункциональная характеристика. Особенности жизненного цикла у разных видов клеток.
Источник: studopedia.org
Хроматин это комплекс макромолекулы ДНК со вспомогательными белками, обеспечивающими ее компактную укладку и стабильность.
Самое удивительное свойство хромосомы это количество ДНК, которое в нее компактно уложено. Человеческий геном это 3х109 пар азотистых оснований в составе нуклеотидов, которые, если полностью развернуть геном одной единственной человеческой клетки, составил бы более метра в длину, но благодаря особой пространственной организации, легко умещается в объем клеточного ядра диаметром 10-5 метра. Это потрясающее инженерное чудо, то, как будучи в составе хромосомы, длинная линейная молекула ДНК, способна обратимо снижать плотность своей укладки – «разматываться», для того чтобы дать возможность необходимым генам экспрессироваться, разделить поровну генетический материал клетки во время митоза (деления клетки) и укладываться компактно вновь.
Источник иллюстрации: micro.magnet.fsu.edu
Структура хроматина зависит от фазы клеточного цикла.
к, во время интерфазы – наиболее продолжительной фазы в клеточном цикле, в ходе которого клетка растет, дифференцируется, синтезирует внутриклеточные компоненты, необходимые для предстоящего митоза такие как РНК и разнообразные белки, происходит репликация (удвоение) ДНК для того чтобы обеспечить генетическим материалом дочернюю клетку. Для синтеза белков в этот период активно используется клеточный генетический материал, поэтому, хроматин будет менее компактным «размотанным», деспирализованным для того, чтобы позволить РНК полимеразам транскрибировать (переписывать с ДНК информацию о первичной структуре белка), такой хроматин содержит большее число активных генов и на светооптическом уровне будет выглядеть более оптически прозрачным и светлым и именоваться эухроматином, таким образом, чем более синтетически активна клетка, тем больше эухроматина будет содержать ее ядро.
Участки ДНК несущие неактивные — «выключенные» гены плотно спирализованные и уложенные в катушки с помощью структурных белков гистонов будут окрашиваться гематоксилином более интенсивно и на светооптическом уровне выглядеть как более темные сине фиолетовые, базофильные участки, или гранулы в ядре клетки.Источник иллюстрации: histology.leeds.ac.uk
По мере того как клетка приближается к моменту вступления в процесс митоза – способ деления соматических клеток организма (не половых) ну, или мейоза в том случае, если это половая клетка, хроматин упаковывается настолько плотно, насколько это вообще возможно, для того чтобы облегчить сегрегацию (разделение) генетического материала материнской клетки от генетического материала дочерней клетки. Во многих клетках такой сверхконденсированный генетический материал, организованный в хромосомы, может быть даже увиден в светооптический микроскоп.
Источник иллюстрации: cytologystuff.com
В опухолевых клетках, как синтетические процессы, так и митотическая активность находятся, как правило, на чрезвычайно высоком уровне, поэтому паттерн хроматина у них, да и размеры ядра будут значительно отличаться от обычных, нормальных клеток, и помимо прочих изменений ядра, таких как увеличение ядерно-цитоплазматического отношения, формы и количества ядер, появления одного или даже нескольких ядрышек, также заметно изменяется как паттерн распределения хроматина, а также и соотношение эу- и гетерохроматина, что является иногда важнейшим признаком, который в совокупности с многими другими, позволяет дифференцировать диспластические и неопластические клетки от нормальных.
Тонкий, мелкодисперсный, нежный (бластный) хроматин.Источник иллюстрации: medical-labs.net
Как в ядрах этих бластных клеток при остром монобластном лейкозе (AML-M5a).
Источник иллюстрации: pathpedia.com
Мелкозернистый хроматин
Источник иллюстрации: medical-labs.net
Как в ядрах этих лимфобластов при Т-лимфобластном лейкозе
Источник иллюстрации: pubcan.org
Кружевной или ретикулярный хроматин
Источник иллюстрации: medical-labs.net
Как в ядрах незрелых клеток из аспирата костного мозга при миеломной болезни (клетки в верхней части снимка), более зрелые клетки будут иметь более грубые зерна хроматина (клетки в среднеи и нижней части снимка).
Источник иллюстрации:pubcan.org
Крупнозернистый грубый хроматин
Источник иллюстрации: medical-labs.net
Как в этих лимфоидных клетках из плеврального выпота при медиастинальной Б-клеточной лимфоме.
Источник иллюстрации: geocities.ws
Скомканный, неоднородный хроматин
В норме, такой хроматин характерен для плазматических клеток
Источник иллюстрации: medical-labs.net
Как в клетках этой мелкоклеточной лимфомы
Источник иллюстрации: pathpedia.com
Смазанный, гиперхромный, плотный хроматин
в норме, такой хроматин характерен для лимфоцитов
Источник иллюстрации: medical-labs.net
Как в клетках этой паратиреоидной аденомы
Источник иллюстрации: webpathology.com
Понятно, что границы между этими подгрупами довольно условны и критериев особенно никаких нет, кроме того, данные паттерны, могут довольно часто сочетаться.
И, на закуску, для сравнения:
Источник иллюстрации: medical-labs.net
Источник: andreas-zarus.livejournal.com
Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК.
При наблюдении некоторых живых клеток, особенно растительных или же клеток после фиксации и окраски, внутри ядра выявляются зоны плотного вещества. В состав хроматина входит ДНК в комплексе с белком. В интерфазных клетках хроматин может равномерно заполнять объем ядра или же располагаться отдельными сгустками (хромоцентры). Часто он особенно четко выявляется на периферии ядра (пристеночный, примембранный хроматин) или образует внутри ядра переплетения довольно толстых (около 0. 3 мкм) и длинных тяжей, образующих подобие внутриядерной цепи.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца (хромосомы), которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (иногда называемого гетерохроматин). Показано, что степень деконденсации хромосомного материала в интерфазе может отражать функциональную нагрузку этой структуры. Чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы. Падение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных телец — хромосом. В этот период хромосомы не несут никаких синтетических нагрузок, в них не происходит включение предшественников ДНК и РНК.
Исходя из этого можно считать, что хромосомы клеток могут находиться в двух структурно-функциональных состояниях:
В рабочем, частично или полностью деконденсированном, когда с их участием в интерфазном ядре происходят процессы транскрипции и редупликации;
В неактивном — в состоянии метаболического покоя при максимальной их конденсированности, когда они выполняют функцию распределения и перенося генетического материала в дочерние клетки.
В химическом отношении препараты хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов, в состав которых входит ДНК и специальные хромосомные белки — гистоны. В составе хроматина обнаружено также РНК. В количественном отношении ДНК, белок и РНК находятся как 1: 1, 3: 0, 2. О значении РНК в составе хроматина еще нет достаточно однозначных данных. Возможно, что эта РНК представляет собой сопутствующую препарату функцию синтезирующейся РНК и поэтому частично связанной с ДНК или это особый вид РНК, характерный для структуры хроматина.
Схема конденсации хроматина:
Источник: biology623.blogspot.com